ConnectedScience:Learning Biology through Constructing and Testing Computational Theories - an Embodied Modeling Approach
|
Uri Wilensky |
Stanford University kreisman@stanford.edu |
Center for
Connected Learning and Computer-Based Modeling
Tufts University
1.0. Introduction
There is a sharp contrast between the picture of the field of biology as studied in school settings and the picture that emerges from the practice of current biology research. While the two pictures are linked by similar content and the objects of study are recognizably the same, the processes involved in the two activities are quite different.
In school settings, typical instruction emphasizes the memorization of classification schemas and established theories. In middle school, classification may take the form of learning the names of the bones of the body, the names and shapes of different plant leaves or the phyla in the animal kingdom. In high school and early undergraduate studies, the content broadens to include unseen phenomena such as parts of the cell or types of protozoa, but the processes of memorizing classifications remains essentially the same. Even in cases where the theories are not yet established, such as the extinction of the dinosaurs, the alternative theories are presented as competing stories to be memorized. And even when students are exposed to research techniques in laboratory work, the emphasis is on following a prescribed procedure rather than reasoning from the evidence gathered in the procedure.
This picture contrasts sharply with the picture that emerges from the recent biology research literature. In this picture, the participants are active theorizers. They devise new evidence gathering methods to test their theories. Instead of accepting classifications as given, they are seen as provisional theories that are constantly reassessed and reconstructed in light of the dialogue between theory and evidence. They reason both forwards, by constructing theories that are consistent with the known evidence and backwards by deducing consequences of theories and searching for confirming/disconfirming evidence. In constructing or assessing an account of a biological phenomenon, they focus on the plausibility of the mechanism proposed — can it achieve the task assigned it in a biologically feasible manner? This assessment of the mechanism often involves reasoning across a range of levels — they ask: is the mechanism constrained by the structure at the molecular, the cellular, the organismic and/or the ecological level?
The contrast between the processes in which these two communities are engaged leads biology students to form a misleading picture of the biological research enterprise. Students form beliefs that biology is a discipline in which observation and classification dominate and reasoning about theories is rare. Furthermore, they believe that learning biology consists of absorbing the theories of experts and that constructing and testing their own theories is out of reach. In this paper, we present an approach that attempts to narrow the gap between school biology and research biology. The approach centers on the use of innovative computer modeling tools that enable students to learn biology through processes of constructing and testing theories.
In recent years, several educational research projects (Jackson et al, 1996; Ogborn, 1999; Roberts & Barclay, 1988) have employed computer modeling tools in science instruction. The approach taken herein differs from these approaches in its use of object-based modeling languages that enable students to model biological elements at the level of the individual (e.g., individual wolf/sheep) as opposed to aggregate (differential-equation based) modeling languages that model at the level of the population (wolf/sheep populations). This technical advance in modeling languages enables students to employ their knowledge of the behavior of individual organisms (or molecules, cells, genes..) in the construction of theories about the behavior of populations of organisms. They embody their theories of individual behavior in a computational agent. This ability to model individual behavior enables students to employ their personal experience with sensing and locomoting in the world as initial elements in their models of other organisms.
In previous work, the authors and other object-based modeling projects (Repenning, 1994; Resnick, 1994; Smith et al, 1994; Wilensky, 1995; 1999; Wilensky & Resnick, 1999) have described the "embodied modeling" approach in a broad inter-disciplinary context. In this paper, we explore the use of this approach, specifically, in biology instruction.
We begin, in the following section, by describing our embodied approach to biological modeling and the object-based parallel modeling language, StarLogoT, in which the models are constructed. In section three, we illustrate this approach by developing embodied models of predator-prey population fluctuations. In section four, we develop a computational model of synchronously flashing fireflies (these species of fireflies are prevalent in the far east, especially Thailand) to frame a discussion of the student modeling process and the relationship of this process to modeling within science. Finally, in our concluding remarks we summarize the major points of the paper. The student modelers described below were participants in the "Connected Mathematics" (Wilensky, 1993; 1995), the ConnectedScience (Wilensky, 1999) and the "Making sense of Complex Phenomena" (MSCP) (Wilensky, 1997; 1999) projects housed at the Center for Connected Learning & Computer-Based Modeling (CCL) at Tufts University. In these projects, students learn about complex systems through construction of object-based parallel models of these systems. The goal of the MSCP project is to construct computational toolkits that enable students to construct models of complex systems and to study students engaged in using these toolkits to model complex systems and to make sense of their behavioral dynamics. Research has documented the difficulties people have in making sense of emergent phenomena, global patterns that arise from distributed interactions, central to the study of complex systems. We have labeled the constellation of difficulties in understanding emergent phenomena and constructing distributed explanations of such phenomena the deterministic/centralized mindset (Resnick & Wilensky, 1993; Wilensky & Resnick, 1995, 1999; Resnick, 1996). In the CCL projects, we have worked with a wide variety of students, ranging from middle school students to graduate student researchers as well as both pre-service and in-service teachers on moving beyond this mindset to a richer understanding of the dynamics of complex systems. Major research sites include two urban Boston high schools. Students from these schools participated in the project as part of their classroom work. Undergraduates and pre-service teachers participated in the context of teacher education courses at Tufts University. Some students participated through informal contexts, pursuing modeling investigations in after-school settings or at the laboratory, housed at the project site. In the classroom context, students, typically, were involved in an extended classroom modeling project led by the classroom teacher and assisted by project researchers. The role of the researchers was to document student work through videotaping and field notes and to support students and teachers in the use of project materials and modeling languages. Such support included the dissemination of interesting cases to be potential sources of models, bringing in books and web sites that might be useful to the modelers. Project researchers also engaged students in structured activities (including participatory simulations not involving the computer (Resnick & Wilensky, 1998)) that would foster reflection on the concept of emergence. They also provided support to students and teachers on the syntax of the modeling language. The computational models described in this paper were built in an object-based parallel modeling language called StarLogoT (Resnick, 1994, 1996; Wilensky, 1995, 1999). In the next section, we describe the workings of StarLogoT and its advantages for modeling biological phenomena.
2.0. The StarLogoT Modeling Language
StarLogoT is a
general-purpose (domain independent) modeling language that
facilitates the modeling of complex systems. It works by providing the
modeler with a framework to represent or "embody" the basic
elements — the smallest constituents — of a system, and then
provides a way to simulate the interactions between these
elements. With StarLogoT, students write rules for hundreds or
thousands of these basic elements specifying how they should behave
and interact with one another. These individual elements are referred
to as ‘turtles’. (StarLogoT owes the ‘turtle’
object to the Logo computer language). Turtles are situated on a two
dimensional grid on which they can move around. Each cell on the grid
is called a ‘patch’, and patches may also execute
instructions and interact with turtles and other patches. Some typical
commands for a turtle are, to move in a given direction, to change
color, to set a variable according to some value, to "hatch"
new turtles, or to look at the properties (variables) of other
turtles. Turtles can also generate random values, so that they can,
for example, execute a sequence of commands with a fixed
probability. Patches can execute similar commands, though they cannot
change location. The wide range of commands executable by turtles and
patches makes it possible to use them to represent many different
systems. For example, turtles can be made to represent molecules,
cells, or individual organisms, while patches represent the medium
(whatever it may be) in which they interact. Time in StarLogoT is
represented as a discrete sequence of ‘clock-ticks’. At each
clock-tick, each turtle and patch is called upon to
execute the rules that have been written for it.
The modeling approach we describe — instantiating the individual elements of a system and simulating their interactions — is not unique to StarLogoT. Such models have been used across a wide variety of domains and have been referred to by several different labels, including: object-based parallel models (Wilensky, 1995; 1997), agent-based models (Beer, 1990; Maes, 1990; Repenning, 1994; Epstein & Axtell, 1996), and individual-based models (Huston, 1988; Judson, 1994). These ‘new wave’ modeling approaches have transformed biology research practice enabling researchers to model increasingly complex multi-leveled biological systems (Forrest, 1989; Langton, 1993; Keen & Spain, 1990; Taylor et al, 1989). For the remainder of this paper, we will employ the term "embodied modeling" to refer to this general approach. While the term, "object-based parallel modeling", which we have used in the past is, perhaps, a more accurate description of the technical workings of StarLogoT, the "embodied modeling" label more closely matches the experience of a biology modeler who is actively engaged in understanding and embodying the behavior of individual biological elements.
In the following two sections of the paper, we will illustrate the embodied modeling approach in biology with two examples of modeling biological phenomena. We intend these examples to illustrate both how such an approach can 1) facilitate the creation of predictive multi-level models in biology and 2) enable biology students to create more powerful explanations of and deepen their understanding of biological phenomena.
3.0. Modeling Predator-Prey Population Dynamics
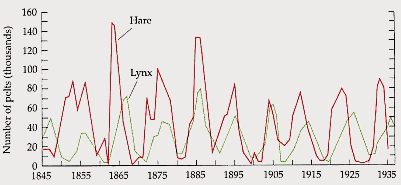
The dynamics of interacting populations of predators and their prey have long been a topic of interest in population biology. Comparisons of a number of case studies have revealed similar dynamics between such populations, regardless of the specific species under study and the details of their interactions. (Elton, 1966). Notably, when the sizes of the predator and prey populations are compared over many generations, we tend to find regular oscillations in these sizes which are out of phase; where one increases, the other tends to decline, and vice-versa (figure 1). In this section, we will present embodied models, developed by students, that reproduce these dynamics.
Figure 1
. Fluctuations of the sizes of predatory lynx and prey hare populations in Northern Canada from 1845-1935 (Purves et al, 1992).3.1. The embodied approach
Using embodied tools, such as StarLogoT, students, typically, first approach the modeling problem by asking what kinds of rules an individual predator or individual prey must follow so that, when allowed to interact in large numbers, populations of such individuals will exhibit population oscillations. It may seem to readers that one would need to be highly familiar with the phenomenon being modeled and with current theories in order to develop a working set of rules, but our experience indicates otherwise. In the Making Sense of Complex Phenomena project, we have found that students are often able to develop solid explanatory models of various phenomena, with only a small amount of background knowledge. We generally encourage modelers to try and make sense of a problem on their own before seeking external resources, and often they are quite surprised at how far they are able to get. To help convey a sense of this process, we will describe the development of a StarLogoT predation model from the standpoint of a student, Talia. While Talia’s case has its own unique characteristics, the topic of predator/prey interactions is popular with high school students and, thus, we have observed many "average" students going through a modeling process quite similar to Talia’s.
3.2. Finding rules for wolves - an initial model
Talia’s task was to formulate a plausible set of rules for a typical predator and a typical prey. Recall that the characteristic properties of predator-prey interactions are invariant across many species and many different conditions. Rather then be specific, then, these rules needed to point to general behaviors that all such species can be said to perform in one way or another. In her first attempt, she described a predator (say, a wolf) as moving about in the StarLogoT world and looking for prey (say, sheep). As the wolf needs energy to live (which it must obtain by eating sheep), she decided that each step in the world will cost the wolf energy. Running out of energy will cause the wolf to die, and the only way to gain energy is by eating sheep. Here is a simple rule-set for a wolf based on the Talia’s description (stated in summary form here):
Rule-set W1: wolf
at each clock-tick:
1. move randomly to an adjacent patch and decrease energy by E1
2. if on the same patch as one or more sheep, then eat a sheep and increase energy by E2
3. if energy 0 then die
4. with probability R1 reproduce
Talia gave the sheep a simpler rule-set. Their job is only to move about and reproduce, though they may still be eaten by the wolves:
Rule-set S1: sheep
at each clock-tick:
1. move randomly to an adjacent patch
2. with a probability of R2, reproduce
3.3. Running the model
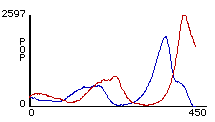
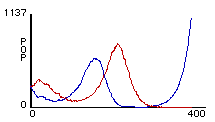
Figures 2a
(left) and 2b (right). Two different outcomes from rule-sets W1 + S1. Red lines represent predator population size and blue lines represents prey population size.After Talia ran her model several times, she noticed that one of two general outcomes would always occur. Either there were oscillations until all the sheep were eaten, whereupon the wolves died from starvation (figure 2a). Or, there were oscillations until the number of sheep dipped too low and the wolves all died off, at which point the sheep reproduced exponentially (figure 2b). Her simple rule-set thus succeeded in producing oscillations, but the pattern was unstable. The next logical step in the modeling process was for her to determine the cause of this instability and correct her model. In order to do this, Talia engaged in a process of successive revision — she would repeatedly devise some variation of her rule-set, and then program it and observe its effects.
3.4. Researching the relevant biological literature
Research into scientific literature is often a part of the model development process. This can help amend any errors in a student’s knowledge of the phenomenon or reveal any important facts that the student might be overlooking. After experiencing difficulty devising a rule-set that would lead to stable oscillations, Talia did some research to determine the source of the problem. Notably, she read that when such systems were first created in the laboratory the findings were very similar to her StarLogoT model: either the predators ate all the prey and then starved, or the predators first died, and then the prey multiplied to the carrying capacity of the environment (Gause, 1934). Several differences between the natural and experimental/model settings could account for this discrepancy. The most significant such factor was the lack of constraints on the growth of the prey population in the experimental settings. In nature, the size and rate of growth of the prey population are constrained by several factors, including limits on the food resources available to prey and limits on their maximum density (Luckinbill, 1973). The laboratory experiments and Talia’s model, however, included abundant food for the prey, and no other adversities in the system but the possibility of predation.
3.5. Revising the model
The major disparity between the experimental setting and the natural case study was the lack of constraints on the growth of the prey population. Talia addressed this by including a third species within her model — grass — which the sheep must feed on, and which is available in limited supply. There were then ways the prey could die — either by being eaten or by starving. This yielded an updated rule-set for sheep and a new rule-set for grass:
Rule-set S2: sheep
at each clock-tick:
1. move randomly to an adjacent patch and decrease energy by E3
2. if on grassy patch, then eat ‘grass’ and increase energy by E4
3. if energy < 0 then die
4. with probability R1 reproduce
Rule-set P1: patches
at each clock-tick:
1. If green, then do nothing
2. If brown, then wait X1 clock-ticks and turn green
Shown in figure 3 is a typical outcome from rule-sets W1 + S2 + P1. This plot shows the population levels of the predators, the prey, and the grass
.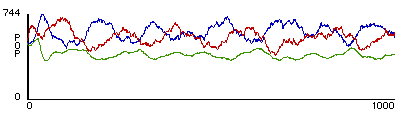
Figure 3
. A typical outcome from rule-sets W1+ S2 + P1. Red represents predator population size, blue represents prey population size, and green represents relative amount of grass.Talia was surprised to find this model much more stable. Oddly enough, she saw that limiting the resources of the prey population actually increased their chances of survival. Conversely, by allowing the prey population unlimited food, they were actually more likely to die off! This surprising result, just as true in the natural world as it is in the StarLogoT world, is known in the scientific literature as the ‘paradox of enrichment’ (Rosenzweig, 1971). Talia was pleased to find that her revised model had correctly predicted this result.
4.0. Modeling Synchronized Fireflies
In this section, we present a second example of how students can use StarLogoT as a laboratory for exploring biological mechanisms. Our example follows the inquiry of an undergraduate student, Paul, a philosophy major whose formal biology instruction consisted solely of high school biology courses. Paul had heard of the phenomena of synchronously flashing fireflies and was intrigued. The following paragraph will provide some background.
For centuries, travelers along Thailand’s Chao Phraya River have returned with stories of the astonishing flashing fireflies that inhabit the mangrove trees along the banks. Come nightfall, the fireflies in a given tree will pulse their lights, on and off, in near perfect synchrony with one another. There are several species of firefly that are known to do this, such as the Southeast Asian Pteroptyx Malacae and Pteroptyx Cribellata. When one such firefly is isolated, it will typically emit flashes at regular intervals. When two or more such fireflies are placed together they entrain to each other — that is, they gradually converge upon the same rhythm, until the group is flashing in synchrony (Buck, 1988).
How do the fireflies achieve this coordinated behavior? When we think about how behavior is coordinated in our daily lives, we tend to think of schedules and elaborate plans. Paul was perplexed at how creatures that seem to have little capacity for intelligent planning are nonetheless capable of such coordination. It was Paul’s suspicion that there must be a simple mechanism behind the feat of the synchronizing fireflies. His goal was to try to understand this mechanism by building a model of it in StarLogoT.
4.1. Approaching the problem — initial assumptions
To begin, Paul made several working assumptions about these fireflies — he was prepared to revise them later if necessary. First, he decided that the mechanism of coordination was almost certainly a distributed mechanism. That is, the fireflies were not all looking to a leader firefly (or some other stimulus) for "flashing orders", but rather were achieving their coordination through passing and/or receiving messages from other fireflies. From his previous experience with StarLogoT, he had learned that not all coordinated group behavior requires a leader to direct the group (see Resnick, 1996; Wilensky & Resnick, 1999). A second assumption, following the first, was that the system could be modeled with only one set of firefly rules; that is, with every firefly in the system following the same set of rules. Although he recognized that this assumption might have been too strong, just as ant and bee populations do divide roles among their groups, he decided to first try out the simpler hypothesis of undifferentiated fireflies. Yet a third assumption Paul made concerned the movement of the fireflies — that it was not necessary to model this movement as coordinated or governed by deterministic rules, but rather it could be modeled as random flights and turns. From experience with other StarLogoT models, he had come to appreciate the role of randomness in enabling coordination (Wilensky, 1997; 1999). In a wide variety of domains, ranging from the movements of particles in a gas, to the schooling of fish and the growth of plant roots, Paul had seen how stable organization could emerge from non-deterministic underlying rules. A final assumption was that the behavior of the fireflies could be modeled in two dimensions.
These assumptions transformed Paul’s task into one of finding a plausible set of rules for a typical firefly. Since Paul knew that a firefly left to its own would simply continue to flash at a regular pace, he reasoned that there were now two principle questions that he had to answer. The first was to determine what would trigger a firefly to change the timing of its flash, and the second was to determine what the response of the firefly would be to this trigger stimulus.
4.2. Thinking like a firefly
Often when building a model, students find it helpful to identify with the individuals within the model and to view phenomenon from their perspective. In order to attack the first of his two questions, Paul began to "think like a firefly". He reasoned along the following lines: If I were a firefly in that situation, what information would I have to go on? It would be dark, and I probably wouldn’t be able to see the other fireflies. I probably wouldn’t have much capacity for hearing or sensing the other fireflies either. I would, however, be able to see their flashes. Perhaps, then, I could look to see who else is flashing and then use this information to adjust my own flashing pattern. From this, Paul provisionally concluded that the trigger stimulus was probably the flashing of other fireflies.
Next, he considered what the response of a firefly would be to this stimulus. Paul could think of several possibilities: a firefly could flash in response, a firefly could increase or decrease the delay until the next flash, or perhaps the firefly could modulate this delay depending on the ‘strength’ of the stimulus. He found it was difficult to intuit the effect of each possibility — none of them result in synchronization in any obvious fashion. The next step was to test each mechanism by coding it up as StarLogoT model and observing the effects.
4.3 Putting it together — an initial model
Paul consolidated his decisions and assembled an initial model to test out various mechanisms. To regulate rhythmic flashing behavior, the model firefly incorporates a timer that continually counts down from a parameter R to zero and then resets to R and begins the cycle again. Each time the timer reaches zero, it causes the firefly to flash — that is, the firefly changes its color to yellow for one clock-tick (rules F1.1 — F1.3). The firefly also ‘flies’ around by moving randomly at each clock-tick (rule F1.4). Finally, rule F1.5 is the flash reset rule. Paul began with a flash reset rule would, in response to any flash seen within an adjacent patch, cause the firefly to flash immediately and reset its timer to R.
Rule-set F1: firefly
to initialize:
0. set timer with random value between 0 and R
at each clock-tick:
1. if color is yellow (flash is on), then change color to black (flash is off)
2. if timer is zero, then change color to yellow and reset timer to R
3. decrement countdown timer by one
4. move randomly to an adjacent patch
5. if there is a yellow firefly within one patch, then change color to yellow (flash is on) and reset the timer to R
When Paul coded up this rule-set in the StarLogoT language and executed it, he found that it didn’t quite work. In fact, it did cause flashing behavior to propagate through the population, but the fireflies couldn’t stop flashing — that is, they perpetually caused each other to flash until the result was one persistent flash. In response, Paul experimented with several other flash reset mechanisms and variations to his rules. Though he observed the emergence of several interesting patterns, he was unable to find a plausible rule-set to give him the precise behavior he was interested in.
4.4. Researching the relevant biological literature
Notice how far Paul was able to get without reference to detailed research on real fireflies. From his initial goal to model ‘whatever’ was going on, he was able to reason to the point where he was seeking a very particular sort of mechanism. At this point, Paul did some research into the scientific literature in order to obtain information about real fireflies (Buck, 1988; Buck, 1976; Buck, 1968). From this research, Paul discovered that many of his design decision were, in fact, biologically plausible. This includes his focus on a distributed synchronization mechanism, his assumption of undifferentiated fireflies, and his conclusion that flashing serves as the means of communication for synchronization. He also learned that the mechanisms of synchronization he had tested out were not far from the actual mechanisms in fireflies. Though different methods of synchronization are seen across species, one strategy that is similar to Paul’s strategy is called ‘phase-delay synchronization’. Under this strategy, when a firefly perceives a flash it delays its next flash so that it will occur one flashing period after the perceived flash. This is the strategy known to be employed by the Southeast Asian fireflies of the Pteroptyx genus. Paul’s next step was to alter his existing model in order to determine whether this strategy would actually work.
4.5. Revising the model
Paul’s rule set needed only slight modification in order to test the phase-delay synchronization strategy. He altered it as follows:
Rule-set F2: firefly
0-4. Identical to rule-set F1
5. if there is a yellow firefly within one patch, then reset the timer to R
Paul executed rule-set F2 using 1000 fireflies, and was amazed to see the model fireflies converge upon a single rhythm before his eyes (figure 4). Paul was encouraged by the initial results of his investigation, and was left with even more questions to consider and to research. For example, he was intrigued by the ability of some fireflies to adapt not only the timing of their flash, but also the duration between flashes. The papers he had looked at gave no complete theory of how this could be done. He was also interested in customizing his model to reflect the idiosyncrasies (e.g. multiple consecutive flashes, responses to irregular stimuli) of particular species, such as Pteropox Malacae and Photinus Pyralis. Though he began his inquiry with only a single question in mind, he found that his questions multiplied as his research continued.
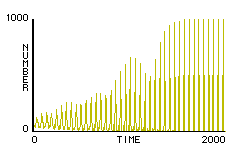
Figure 4. Typical plot of the number of flashes in a population at a given time under rule-set F2.
5.0. Concluding Remarks
The embodied modeling approach we have presented and illustrated herein can be the basis of a modeling-centered biology curriculum in secondary and post-secondary contexts. By removing the barriers of formal mathematical requirements, it enables students to meaningfully engage the dynamics of complex biological systems. They are able to construct models of such systems, reason about the mechanisms that underlie them and predict their future behavior. Because they are able to use their knowledge of the individual elements in the system to construct their model, they are provided with an incremental path to constructing robust models. When their knowledge of the individual biological elements is combined with their knowledge of their own embodiment, their own point of view, they are enabled to think like a wolf, a sheep or a firefly.
The above examples have, we hope, demonstrated the power of the embodied modeling approach to enable students to construct robust models and engage in exciting scientific inquiry. The modeling-based classroom is dramatically different from most venues of classroom practice. Rather than passively receiving an authority’s explanation of science and mathematics concepts, students seek out and consider these concepts on their own. Rather than carry out the directions for predetermined lab studies, students engage in new investigations. What underlies this approach is our deep conviction of the value of reasoning about scientific order. In both the predation and firefly examples presented above, students were encouraged to reason through a problem, creating and testing their own theories and hypotheses, before reaching for the established literature.
There are, however, significant practical obstacles to employing the embodied modeling approach on a large scale at this point in time. Foremost amongst these is the unfamiliarity, to most teachers, of the modes of operation of the modeling-based classroom as described above. Overcoming this obstacle will necessitate significant teacher education efforts on the changed role of the teacher in the classroom. Another significant obstacle is the relative sparseness of personal computers in classrooms. StarLogoT requires the computational power of typical current personal computers. To take full advantage of the approach, a class needs to have at least one of these per three students. Many classrooms do not meet this requirement. Finally, the modeling approach requires teachers and students to be comfortable with algorithmic thinking and debugging, topics that are optimally learned through longitudinal developmental strands. As of now, there is not significant commitment to these topics and strands in K-16 education.
Despite these significant obstacles, there are good reasons to believe that the approach we describe will come to be widely incorporated into the science and social science curriculum. Most of the obstacles described above are short-term obstacles. The eventual proliferation of computers in classrooms combined with educational studies describing the effectiveness of the modeling-based classroom and the overwhelming adoption of modeling tools by practicing scientists will put pressure on teacher education programs to adopt these new methods and tools. Indeed, it is already happening in school districts and schools of education that are located near high technology centers.
Our approach promotes several processes of reasoning that are central to science: developing original hypotheses, formalizing ideas, researching existing solutions, and critical analysis of results. We believe that experience with these processes will be of significant advantage to all students as they seek to understand science and, more generally, the world around them. Few students will go on to become scientists. To the ones that don’t, we owe them more than an introductory glimpse of current theories — we owe them the tools with which to appreciate scientific evidence and to reason in a thorough, scientific manner on their own. For the ones that do, we owe them a framework within which they will be better prepared to absorb and appreciate the myriad facts they will encounter for years to come. Thus, it is our hope that the approach we are developing will serve as a framework for all students to be able to work and think like scientists.
Acknowledgments
The preparation of this paper was supported by the National Science Foundation (Grants RED-9552950, REC-9632612). The ideas expressed here do not necessarily reflect the positions of the supporting agency. Rob Froemke played a significant role in the development of the StarLogoT modeling environment with the assistance of Eamon Mckenzie, Paul Deeds and Dan Cozza. Mitchel Resnick, Brian Silverman and Andy Begel were largely responsible for a previous StarLogo version, StarLogo 2.0, on which StarLogoT was based. Steve Longenecker and Josh Mitteldorf contributed greatly to our thinking about the predator-prey models. Ed Hazzard, Walter Stroup and Rob Froemke made valuable clarifying suggestions. We would like to thank Seymour Papert for his overall support and inspiration and for his constructive criticism of this research in its early stages. Mitchel Resnick has been an invaluable collaborator in thinking about issues of complexity and education in general and in exploring the predator prey models in particular. Daniel Dennett provided valuable criticism of the firefly model and inspired many of our observations about embodied modeling.
References
Beer, R.D. (1990). Intelligence as adaptive behavior: An experiment in computational neuroethology. Cambridge, Ma. Academic Press.
Buck, J. (1988) Synchronous Rhythmic Flashing of Fireflies II. Quarterly Review of Biology, 63, 265-287.
Buck, J. & Buck, E. (1976). Synchronous Fireflies. Scientific American, 234, 74-85.
Buck, J. & Buck, E. (1968). Mechanism of Rhythmic Synchronous Flashing of Fireflies. Science, 159, 1319-1327.
Elton, C. (1966). Animal Ecology. New York: October House.
Epstein, J. & Axtell, R. (1996). Growing Artificial Societies: Social Science from the Bottom Up. Washington: Brookings Institution Press.
Forrest, S. (1989). Emergent Computation: self-organizing, collective, and cooperative phenomena in natural and competing networks. Amsterdam: North Holland Pub. Co.
Gause, G.F. (1934). The Struggle for Existence. Baltimore, MD: Williams and Wilkins.
Hammer, D. (1994). Epistemological beliefs in introductory physics, Cognition and Instruction, 12 (2), 151-183.
Hudson, G. (1918). Concerted Flashing of Fireflies. Science, 48, 573-575.
Huffaker, C. (1958). Experimental Studies on Predation: Dispersion Factors and Predator-Prey Oscillations. Hilgardia, 27, 343-383.
Huston, M., DeAngelis, D., Post, W. (1988). New Computer Models Unify Ecological Theory. BioScience, 38 (10), 682-691.
Jackson, S., Stratford, S., Krajcik, J., & Soloway, E. (1996). A Learner-Centered Tool for Students Building Models. Communications of the ACM, 39 (4), 48-49.
Judson, O. (1994). The Rise of the Individual-based Model in Ecology. TREE, 9 (1), 9-14.
Keen, R. & Spain, J (1992). Computer Simulation in Biology. New York, Wiley-Liss.
Langton, C. (1993). Artificial Life III. Reading, Ma: Addison Wesley.
Langton, C. & Burkhardt, G. (1997). Swarm. Santa Fe Institute, Santa Fe: NM.
Luckinbill, L. (1973). Coexistence in Laboratory Populations of Paramecium Aurelia and its Predator Didnium Nasutum. Ecology, 54 (6), 1320-1327.
Maes, P. (1990). Designing Autonomous agents, theory and practice from biology to engineering and back. Cambridge, Ma: MIT Press.
Morse, E. (1916). Fireflies Flashing in Unison. Science, 43, 169-170.
Ogborn, J. (1999). Modeling Clay for Thinking and Learning. In Roberts, N., Feurzeig, W. & Hunter, B. (Eds.) Computer Modeling and Simulation in Science Education. Berlin: Springer Verlag.
Papert, S. (1980). Mindstorms: Children, Computers, and Powerful Ideas. New York: Basic Books.
Purves, W. G, Orians. H, Heller. (1992). Life: The Science of Biology. Salt Lake City, Utah : W.H. Freeman.
Reisman, K. (1998). Distributed Computer-Based Modeling. Unpublished undergraduate honors thesis. Tufts University.
Repenning, A. 1994. Programming substrates to create interactive learning environments. Interactive Llearning Eenvironments, 4 (1), 45-74.
Resnick, M. (1994). Turtles, Termites and Traffic Jams: Explorations in Massively Parallel Microworlds. Cambridge, MA: MIT Press.
Resnick, M. (1996). Beyond the Centralized Mindset. Journal of the Learning Sciences, 5 (1), 1-22.
Resnick, M., & Wilensky, U. (1998). Diving into Complexity: Developing Probabilistic Decentralized Thinking Through Role-Playing Activities. Journal of the Learning Sciences, 7 (2), 153-171.
Roberts, N. & Barclay, T. (1988). Teaching model building to high school students: theory and reality. Journal of Computers in Mathematics and Science Teaching. Fall: 13 - 24.
Rosenzweig, M. (1971). Paradox of Enrichment: Destabilization of Exploitation Ecosystems in Ecological Time. Science, 171, 385-387.
Smith, D. C., Cypher, A., & Spohrer, J. (1994). Kidsim: Programming agents without a programming language. Communications of the ACM, 37 (7), 55-67.
Smith, D. C., Cypher, A., & Spohrer, J. (1994). KidSim: Programming agents without a programming language. Communications of the ACM, 37 (7), 55-67.
Stroup, W. (1996). Embodying a Nominalist Constructivism: Making Graphical Sense of Learning the Calculus of How Much and How Fast. Unpublished Doctoral Dissertation, Cambridge, MA: Harvard Graduate School of Education.
Taylor, C, Jefferson, D, Turner, S & Goldman, S. (1989). RAM: Artifical Life for the exploration of complex biological systems. In C. Langton, (Ed.), Artifical Life. Reading, Ma: Addison Wesley.
Wilensky, U. & Resnick, M. (1999). Thinking in Levels: A Dynamic Systems Perspective to Making Sense of the World. Journal of Science Education and Technology. Vol. 8 No. 1.
Wilensky, U. (1999). GasLab–an Extensible Modeling Toolkit for Exploring Micro- and Macro- Views of Gases. In Roberts, N., Feurzeig, W. & Hunter, B. (Eds.) Computer Modeling and Simulation in Science Education. Berlin: Springer Verlag
Wilensky, U. (1997). What is Normal Anyway? Therapy for Epistemological Anxiety. Educational Studies in Mathematics. Special Edition on Computational Environments in Mathematics Education. Noss R. (Ed.) 33 (2), 171-202.
Wilensky, U. (1996). Modeling Rugby: Kick First, Generalize Later? International Journal of Computers for Mathematical Learning. 1 (1), 125-131.
Wilensky, U. (1995). Learning Probability through Building Computational Models. Proceedings of the Nineteenth International Conference on the Psychology of Mathematics Education. Recife, Brazil, July 1995.
Wilensky, U. (1993). Connected Mathematics: Building Concrete Relationships with Mathematical Knowledge. Doctoral dissertation, Cambridge, MA: Media Laboratory, MIT.